МИНИСТЕРСТВО ОБРАЗОВАНИЯ И НАУКИ РОССИЙСКОЙ ФЕДЕРАЦИИ
ГОСУДАРСТВЕННОЕ ОБРАЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ
ВЫСШЕГО ПРОФЕССИОНАЛЬНОГО ОБРАЗОВАНИЯ
"САМАРСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ"
Кафедра зоологии, генетики и общей экологии
Э.Д. Владимирова
ПСИХОЛОГИЯ ЖИВОТНЫХ
Утверждено редакционно-издательским советом
университета в качестве учебного пособия
Самара
Издательство "Самарский университет"
2010
Рецензенты:
кандидат биологических наук, доцент СГПУ С.И. Павлов, доктор психологических наук К.С. Лисецкий
Программа курса "Зоопсихология"
для студентов 3 курса дневного отделения
биологического факультета
Учебная литература
- --
Годфруа Ж. Что такое психология. В 2-х т. М.: Мир, 1992.
- -- Дьюсбери Д. Поведение животных. Сравнительные аспекты. М.: Мир, 1981.
- -- Зорина З.А., Полетаева И.И. Зоопсихология, элементарное мышление животных. М.: Аспект-Пресс, 2000.
- -- Зорина З.А., Полетаева И.И., Резникова Ж.И. Основы этологии и генетики поведения. М.: Высшая школа, 2002.
- -- Иванов А.А. Этология с основами зоопсихологии. СПб.: Лань, 2007.
- -- Мак-Фарленд Д. Поведение животных. Психобиология, этология и эволюция. М.: Мир, 1988.
- -- Меннинг О. Поведение животных. Вводный курс. М.: Мир, 1982.
- -- Правоторов Г.В. Зоопсихология для гуманитариев. Новосибирск: ЮКАЭА, 2001.
- -- Резникова Ж.И. Интеллект и язык животных и человека. Основы когнитивной экологии. М.: Академкнига, 2005.
- -- Скопичев В.Г. Поведение животных. СПб: Лань, 2009.
- -- Фабри К.Э. Основы зоопсихологии. М.: МГУ, 1993.
- -- Филиппова Г.Г. Зоопсихология и сравнительная психология. М.: Академия, 2004.
- -- Хайнд Р. Поведение животных. Синтез этологии и сравнительной психологии. М.: Мир, 1975.
- -- Хрестоматия по зоопсихологии и сравнительной психологии / Под ред. Н.Н. Мешковой, Е.Ю.Федорович. М.: РПО, 1997.
- -- Шовен Р. Поведение животных. М.: Мир, 1972.
- -- Дьюсбери Д. Поведение животных. Сравнительные аспекты. М.: Мир, 1981.
- -- Предмет, методы и задачи зоопсихологии
2. Методы зоопсихологии - наблюдение и эксперимент
, (1)
- факторы, определяющие поведение,
- вес фактора.
), возрастной (
) и половой (
) принадлежности, физиологического (
) и мотивационного (
) состояния, реализуемой в данный момент времени формы функциональной активности (
). Также подлежит учету комплекс биосоциальных отношений, сложившихся в популяции: (
), (
). Следует, кроме того, принимать во внимание постоянно меняющееся взаимодействие пороговых и подпороговых стимулов (
), действующих на особь или группу в разных модальностях восприятия: оптической, акустической, ольфакторной, тактильной. Непрерывный поток поведения животных должен быть формализован в этологической модели так, чтобы появилась возможность обобщать и сравнивать данные полевых наблюдений, полученные в разное время (суточная и сезонная активности) - (
), (
), а также данные, полученные в различных биотопах (условия микростаций, уровень антропогенной нагрузки) - (
), (
).
в одномерной регрессионной модели показывает, насколько интенсивно изменяется поведение по отношению к изменению данного фактора и трактуется как "коэффициент чувствительности Y к X" (Пузаченко, 2004, с. 133) и представляет собой первую производную:
.

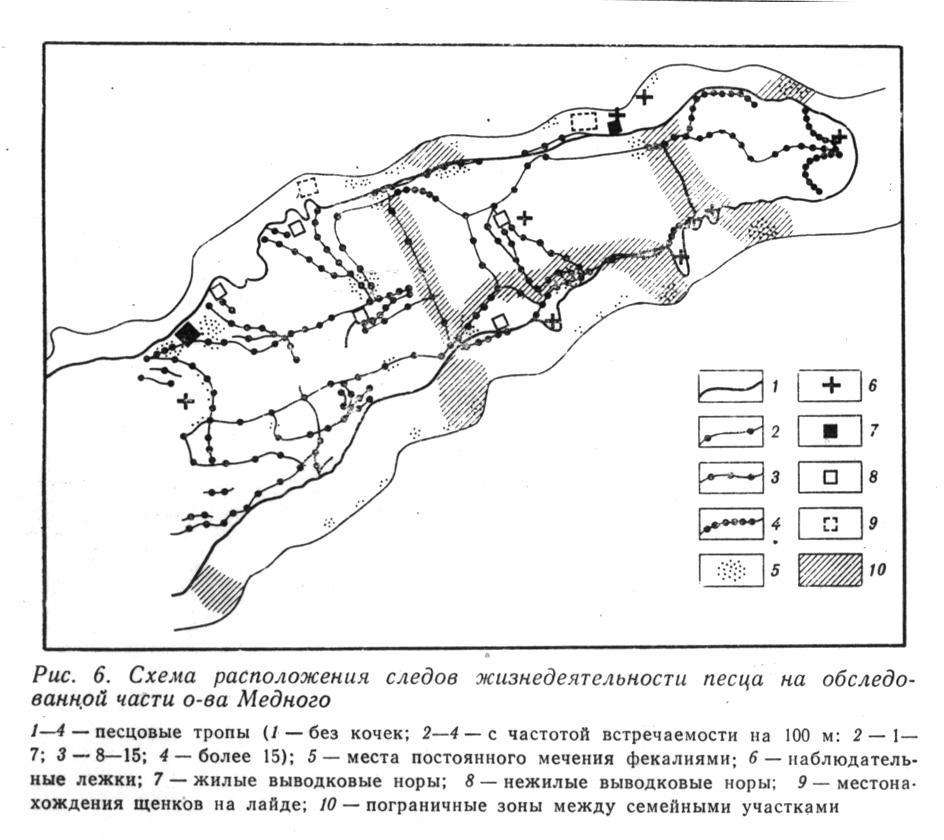
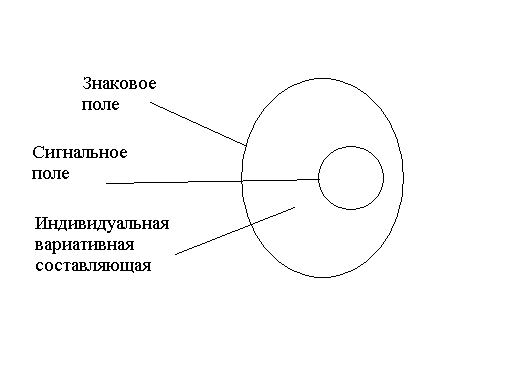
3. Критерии наличия психики
4. Предметы зоопсихологии и этологии. Связь этологии с другими биологическими науками. Экологический подход
к зоопсихологии и этологии
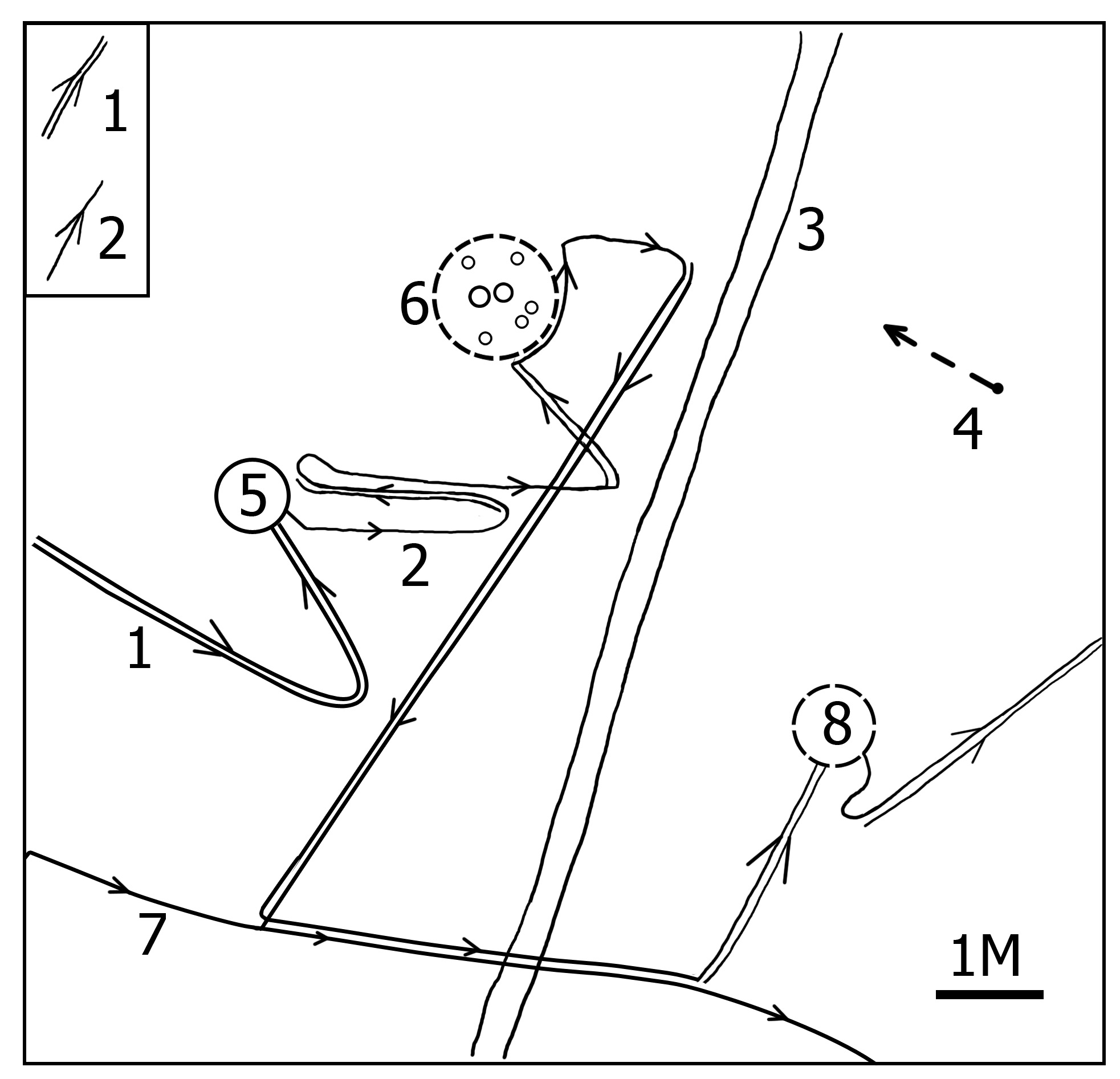
5. Видовой стереотип поведения и лабильные
поведенческие реакции. Функциональные типы активности
6. Проблема антропоморфизма
7. Бихевиоризм
(2)
- сигнальные стимулы первого рода,
- сигнальные стимулы второго рода и т.д.