Specific Functional Forms of Behaviour in Pine marten (Martes martes ),
based on snow tracking data
Видоспецифические функциональные формы поведения лесной куницы (по материалам зимних троплений)
Э.Д. Владимирова
Самарский государственный аэрокосмический университет
имени академика С.П. Королева
Elina J. Vladimirova [elyna-well@nm.ru]
Samara State Aerospace University named after academician S.P. Korolyov
443086, Russia, Samara, Moskovskoye Shosse, 34
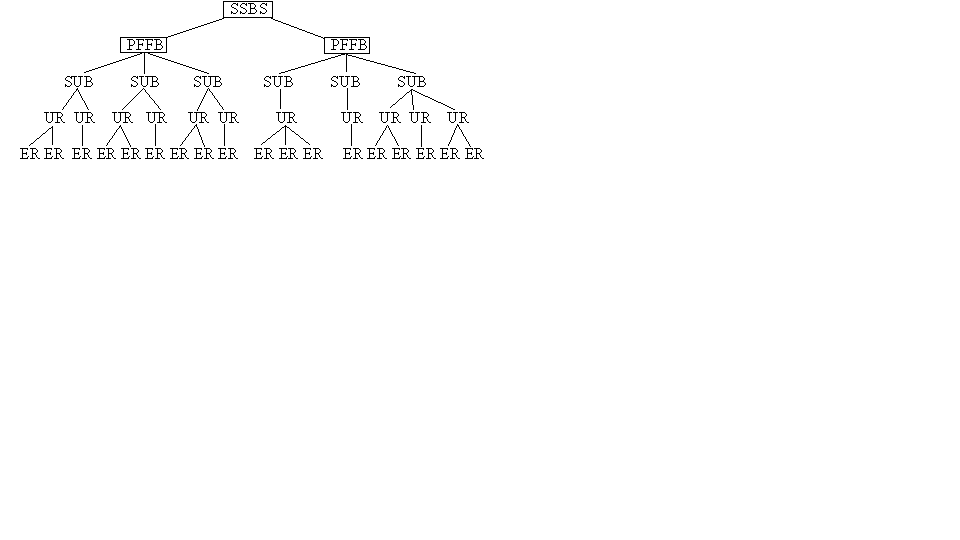
,
, A = {A1, A2, ..., Am},
, B = {B1, B2, ..., Bm},
, C = {C1, C2, ..., Cm},
, D = {D1, D2, ..., Dm},
, где

Таблица
Основные функциональные формы активности лесной куницы
и соответствующие им разновидности двигательных реакций
|
Функциональные формы активности |
в сумме составляющие более 75 % реакций данной функциональной формы |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|